



Why do some plants, like corn or sorghum, grow so vigorously while others appear limited in their vigor and growth under similar environmental conditions?
Our various agronomic plants fit into basic physiological groupings, based upon their varying capacities to perform the ultimate biochemistry, the harvest and processing of sunlight and carbon dioxide into biologically viable carbon and energy through the process of photosynthesis. The following table summarizes key divisions of plants and their relative photosynthetic efficiencies:
Table 1: Physiological grouping of plants by photosynthetic capacity.
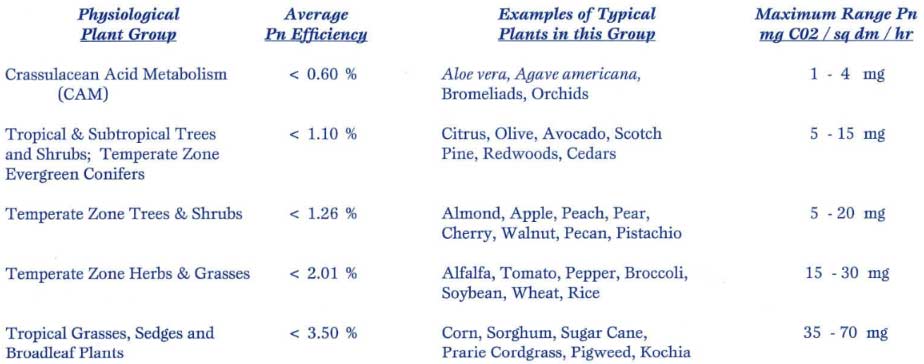 Key: Pn = photosynthesis, Pn Efficiency = Actual Pn energy harvested per energy presented, dm = decimeter (approx. 4” x 4” square), mg = milligram
Key: Pn = photosynthesis, Pn Efficiency = Actual Pn energy harvested per energy presented, dm = decimeter (approx. 4” x 4” square), mg = milligram
As a contrasting example, what is the underlying reason that corn or sorghum perform so much more efficiently than almond or peach?
Without getting into too much detail, one of the main reasons for this large gap in photosynthetic efficiency resides in the difference between the two main types of physiology, ‘carbon-3 physiology’ (C-3) and ‘carbon-4 physiology” (C-4), and how they operate under different conditions.
Figure 1: The Basic Differences Between C-3 and C-4 Physiology
Scenario 1 – Mild Temperatures and Sunlight (generally temperatures between 70 to 95 degrees F)
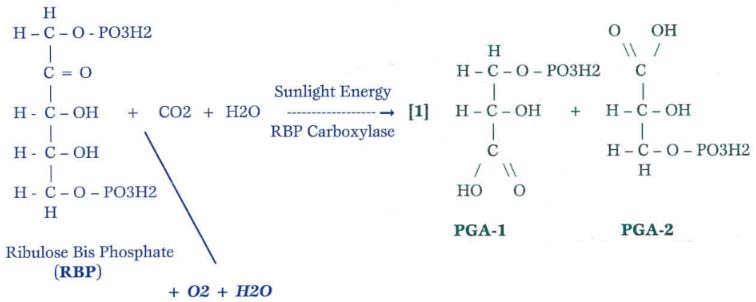 Key: PGA = phosphoglyceric acid
Key: PGA = phosphoglyceric acid
PGA molecules are channeled to various reactions, forming carbohydrates, amino acids, nucleic acids, proteins, cellulose, hemicelluloses, plant hormones, and other positive plant molecules.
Scenario 2 – Hot Temperatures and Intense Sunlight (generally temperatures above 95 degrees F or during physiological shock in which a day’s high temperature exceed a 10 to 15-degree transition from the previous day’s high)
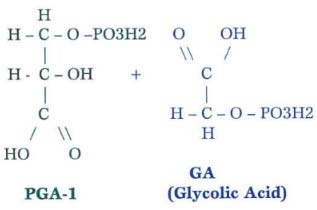 While phosphoglyceric acid can be directly channeled to positive molecules, glycolic acid must undergo a series of energy-expensive reactions before it can be used by the plant. Thus, Scenario 2 results in a more than 50% cut in production. This heat-related impact on photosynthetic efficiency is termed ‘photorespiration,’ also referred to as the ‘Warburg effect.’
While phosphoglyceric acid can be directly channeled to positive molecules, glycolic acid must undergo a series of energy-expensive reactions before it can be used by the plant. Thus, Scenario 2 results in a more than 50% cut in production. This heat-related impact on photosynthetic efficiency is termed ‘photorespiration,’ also referred to as the ‘Warburg effect.’
The more efficient photosynthetic pathway (Scenario 1) is indicative of corn and sorghum, which fall into the C-4 group of plants. Almond and peach host the less efficient photosynthetic pathway (Scenario 2), which fall into the C-3 group of plants. In the hot Central San Joaquin or Sacramento Valley summers, for example, 95+ degree temperatures are a normal occurrence. Furthermore, more than 95% of all agronomic crop species are C-3 plants.
Given the above, it is confusing as to why such an inefficient process would be retained in the genetics of a plant. Why would photorespiration develop and still be retained in C-3 plants today?
As plant life made the transition from small aquatic species to the terrestrial environment and entered the Mesozoic Era—which spanned 225 to 65 million years ago—there were major transitions in the development of plants. Basically, small aquatic plants became adapted to the terrestrial environment. In this new setting, they had an opportunity to diversify and develop multiple physical and physiological forms.
With the prehistoric atmosphere very high in carbon dioxide, land plants flourished, with many species reaching unimaginable size and mass. This tendency towards gigantism also explains the development of the dinosaurs and other large species which dominated the era. However, there is a principle in biology which states that as an organism develops in size and mass, a critical volume is reached beyond which physiological and physical efficiency decline dramatically. In the case of prehistoric plants, there was the need for a check and balance system to limit size and mass and stay within the bounds of operative efficiency. For plants this system turned out to be photorespiration. And while we may perceive photorespiration as a defect, it is indeed a transitional characteristic that occurred for the benefit of the plant kingdom.
Because the C-3 phenomenon of photorespiration is a major stumbling block for plant growers, are there ways to manipulate C-3 plants to minimize the onset of photorespiration and increase crop yields and quality?
While genetic manipulation may be impractical and futuristic, a series of tangible approaches have been explored with success. The technologies explored are based on several principles of plant physiology. First, the basic equation for photosynthetic harvest by a green plant is:
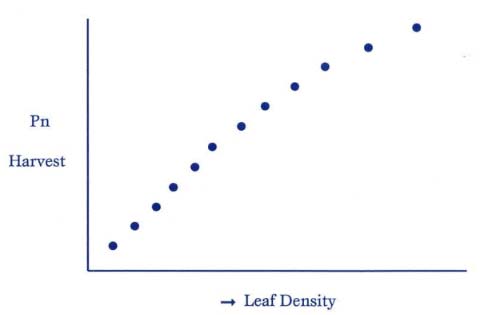
![]() That is, solar energy and the photosynthetic hours per day are basically constant, while a leaf’s surface area and efficiency can be manipulated to maximize crop yields. As it turns out, there is a near straight-line relationship between leaf density and photosynthetic harvest:
That is, solar energy and the photosynthetic hours per day are basically constant, while a leaf’s surface area and efficiency can be manipulated to maximize crop yields. As it turns out, there is a near straight-line relationship between leaf density and photosynthetic harvest:
Figure 2: Relationship Between Leaf Density vs. Photosynthetic Harvest
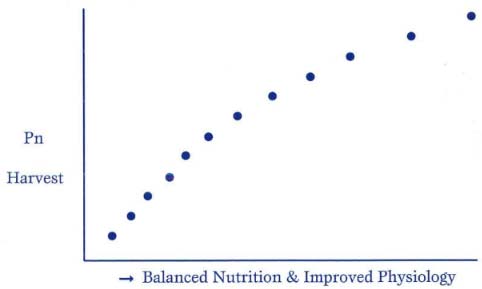
 There is also a parabolic relationship between balanced nutrition and photosynthetic harvest.
There is also a parabolic relationship between balanced nutrition and photosynthetic harvest.
Figure 3: Relationship Between Balanced Nutrition vs. Photosynthetic Harvest
 Thus, when we combine the above three factors, we can begin to draw the blueprints for mitigating photorespiration. The most important factors to any such strategy are balancing the soil and mineral nutrition, using selective microbial enrichment to encourage and/or control the release of minerals and nitrogen, balanced foliar sprays with special emphasis on calcium, potassium, boron, phosphorus, and various carbon compounds. Ideally, these are all applied ahead of or during heat waves. Judicious attention to the details outlined can increase leaf area, density, and increase photosynthetic efficiency, the combined efforts of which create increase tolerance and/or resistance to photorespiration.
Thus, when we combine the above three factors, we can begin to draw the blueprints for mitigating photorespiration. The most important factors to any such strategy are balancing the soil and mineral nutrition, using selective microbial enrichment to encourage and/or control the release of minerals and nitrogen, balanced foliar sprays with special emphasis on calcium, potassium, boron, phosphorus, and various carbon compounds. Ideally, these are all applied ahead of or during heat waves. Judicious attention to the details outlined can increase leaf area, density, and increase photosynthetic efficiency, the combined efforts of which create increase tolerance and/or resistance to photorespiration.
Why do particularly healthy plants seem to be more tolerant or resistant to pathogens and pests?
The most important principle of resistance to pathogens and pests is understanding that it is a rate-related phenomenon. That is, the ability of the host to respond quickly to literally out-race and out-compete the growth and establishment of the intruder is critical:
Figure 4: The Susceptible Host Response vs. Resistant Host Response as a Function of Metabolic Rate
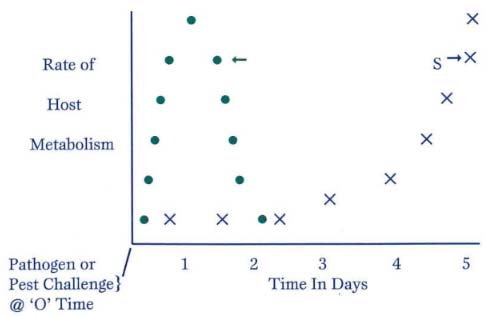 Figure 4 depicts the typical physiological response in hosts that are resistant (the dots) and susceptible (the Xs). By the time the susceptible host recognizes and responds to the intruder, it is too late—the pest or pathogen has already had enough time to damage the host and establish itself. The resistant host immediately recognizes and responds to the pathogen or pest, producing combative compounds within 48 hours to ward off the establishment of the intruder. There is another principle that coincides with the above principle of rate-related resistance:
Figure 4 depicts the typical physiological response in hosts that are resistant (the dots) and susceptible (the Xs). By the time the susceptible host recognizes and responds to the intruder, it is too late—the pest or pathogen has already had enough time to damage the host and establish itself. The resistant host immediately recognizes and responds to the pathogen or pest, producing combative compounds within 48 hours to ward off the establishment of the intruder. There is another principle that coincides with the above principle of rate-related resistance:
Figure 5: The Status of Carbon and Energy Harvest in the Host vs. Resistance and Tolerance to Pests and Pathogens
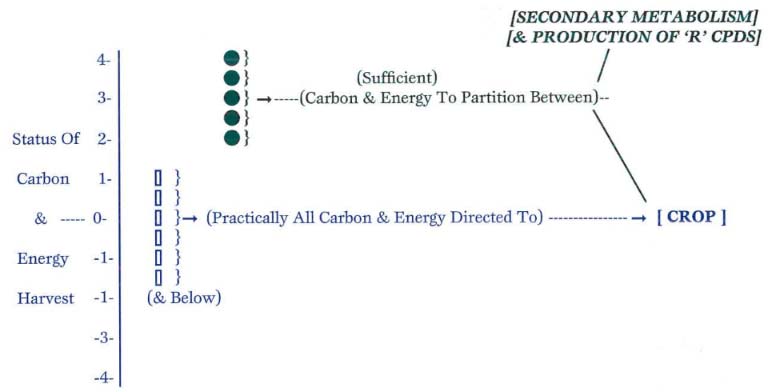 If the plant is operating at a level where the amount of carbon and energy harvested are marginally supplying the needs of the plant (near the ‘0’ point at which carbon and energy harvest barely match the rate of use, also called the ‘compensation point’), then that carbon and energy will be directed almost totally to the needs of the crop. This is the first priority of the plant: to perpetuate its kind through production of reproductive tissues. Conversely, if the plant is operating quite efficiently, at relative health ratings greater than ‘2,’ there will be supplementary carbon and energy. The surplus amounts can be diverted to various other reactions, including the production of secondary metabolites, which are instrumental in imparting the resistance response to pests and pathogens.
If the plant is operating at a level where the amount of carbon and energy harvested are marginally supplying the needs of the plant (near the ‘0’ point at which carbon and energy harvest barely match the rate of use, also called the ‘compensation point’), then that carbon and energy will be directed almost totally to the needs of the crop. This is the first priority of the plant: to perpetuate its kind through production of reproductive tissues. Conversely, if the plant is operating quite efficiently, at relative health ratings greater than ‘2,’ there will be supplementary carbon and energy. The surplus amounts can be diverted to various other reactions, including the production of secondary metabolites, which are instrumental in imparting the resistance response to pests and pathogens.
(Thumbnail source: Publicdomainpictures.net, public domain via CC0 1.0)
